Наиболее распространенный аллель, или, как его первоначально называли, аллель дикого типа, который обычно бывает доминантным для всех остальных аллелей, обозначают либо заглавной буквой (единственный среди других аллелей), либо индексом + (В , с , е , st ). Изредка в природе встречаются доминантные мутации – они обычно обозначаются сочетанием букв, начиная с прописной (например, мутация Bar доминантна по отношению к аллелю дикого типа В ).
При рассмотрении сцепленных генов используют знак "/". При этом, если аллели двух рассматриваемых генов находятся на одной хромосоме, говорят о цис-положении (АВ/ав), а если на разных – о транс-положении (Ав/аВ).
Применяют и специальные символы:
Р – родители;
G – гаметы;
F – поколения (F 1 , F 2, F 3 );
х – знак скрещивания;
♂ – знак мужского пола;
♀ – знак женского пола.
4.2. Генетический анализ
Совокупность методов изучения наследственности получила название "генетический анализ". Его основа – гибридологический метод, разработанный Г. Менделем. С открытия законов наследования Г. Менделем и начинается история генетики. Не меньшая заслуга в становлении этой науки принадлежит Т. Моргану и его школе. Можно считать, что эти ученые заложили фундамент генетики как науки.
Гибридологический метод, разработанный Г. Менделем, показал, что родительские черты не смешиваются в потомстве, а передаются как независимые признаки. В литературе результаты работ Г. Менделя получили названия "законов", хотя он сам не выдвигал их четких формулировок.
1-й закон – закон единообразия гибридов первого поколения или закон доминирования. При скрещивании чистых линий все потомство первого поколения единообразно по исследуемому признаку. Признак, который проявлялся у потомков, стали называть доминантным , признак, который не проявлялся, – рецессивным.
2-й закон – закон расщепления гибридов во втором поколении. Во втором поколении соотношение частоты проявления доминантного и рецессивного признаков составляет 3: 1.
3-й закон – закон независимого наследования признаков. Расщепление по каждой паре признаков идет независимо от других пар признаков.
1-й и 2-й законы Менделя были выведены при скрещивании организмов, различающихся по одному признаку (моногибридное скрещивание), а 3-й закон – по нескольким признакам (полигибридное скрещивание).
В современном понимании законы Г. Менделя показывают случайный характер распределения аллелей в гаметы во время гаметогенеза и их объединения при оплодотворении. Биологический механизм мейоза дает объяснение открытым Г. Менделем закономерностям. Во время мейоза аллели, определяющие альтернативные признаки, расходятся в разные половые клетки, поэтому каждая гамета имеет только один аллель ("правило чистоты гамет"). При оплодотворении происходит объединение гаплоидных хромосомных наборов, поэтому каждый ген в организме представлен в двух вариантах – в отцовской и материнской хромосоме.
Заслуга Г. Менделя проявилась в том, что он выдвинул математически обоснованную и проверяемую гипотезу наследования признаков. Объективно оценить, насколько полученный результат соответствует проверяемой гипотезе, позволяет статистика. В настоящее время сформировалась особая наука – биометрия, занимающаяся математической обработкой биологических данных. Современная генетика наиболее тесно из биологических наук интегрирована с биометрией.
Для проверки истинности своей гипотезы Г. Мендель применил так называемое анализирующее скрещивание доминантной и рецессивной форм. Рецессивная форма (всегда гомозигота) как бы "анализирует" по соотношению потомства генотип формы с доминантным признаком, которая может быть и гомозиготой, и гетерозиготой.
Анализ различных генотипических классов при гибридологическом анализе облегчает пользование решеткой Пеннета (рис. 4.1). Р. Пеннет (1875–1967) – известный английский генетик, впервые предложивший этот метод для гибридологического анализа. Ему же принадлежит термин "менделизм".
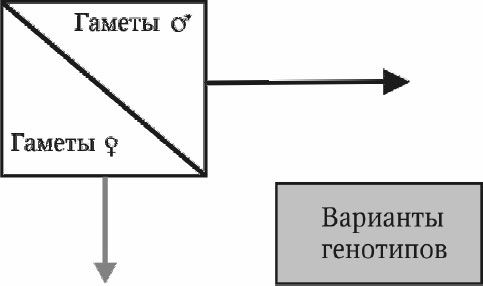
Рис. 4.1. Решетка Пеннета
Число возможных вариантов гамет, генотипов и фенотипов легко рассчитать по специальным формулам, в которых n – число гетерозиготных локусов:
– число вариантов гамет – 2 ;
– число вариантов генотипов – 3 ;
– число вариантов фенотипов при полном доминировании – 2 .
В результате работ американского генетика Т. Моргана и его школы сформировалась хромосомная теория наследственности, суть которой состоит в следующем:
1. Гены располагаются в хромосомах в линейной последовательности.
2. Каждая хромосома представляет группу сцепленных генов.
3. Каждый ген занимает в хромосоме определенное место – локус.
Локус – это участок расположения гена на хромосоме. Хромосомы содержат последовательности генных локусов, причем у гомологичных хромосом эти последовательности одинаковые.
Поскольку число генов в организме несоизмеримо больше числа хромосом, понятно, что каждая хромосома любого организма несет много генов. Гены, расположенные на одной хромосоме, являются сцепленными . Аллели сцепленных генов наследуются совместно.
Однако сцепление не является абсолютным. В результате кроссинговера сцепленные гены могут быть разъединены и при мейотическом делении они оказываются в разных гаметах. Такие гаметы называются кроссоверными. Поскольку кроссинговер является обязательным процессом и происходит в каждой паре гомологичных хромосом, А. Стертевант выдвинул гипотезу, что частота кроссинговера на участке между генами, локализованными на одной хромосоме, может служить мерой расстояния, на котором они находятся друг от друга. Это предположение подтвердилось. Чем дальше расположены друг от друга гены на хромосоме, тем выше вероятность кроссинговера между ними. Показатели частоты кроссинговера стали использовать для определения расположения генов на хромосоме и составления генетических карт, что стало одним из ведущих направлений генетического анализа.
4.3. Взаимодействие генов
В организме одновременно функционирует множество генов. В процессах реализации генетической информации в признак возможны многочисленные "пункты" взаимодействия разных генов на уровне биохимических реакций. Такие взаимодействия неизбежно отражаются на формировании фенотипа.
Аллельные гены определяют альтернативные признаки, так как лежат в гомологичных локусах гомологичных хромосом. Между аллелями гетерозиготы возможны определенные взаимодействия, лежащие в основе проявления признака в фенотипе. Известны три основные формы межаллельных взаимодействий.
Полное доминирование – в гетерозиготе один аллель (доминантный) подавляет проявление второго аллеля (рецессивного).
Неполное доминирование – в гетерозиготе наблюдается промежуточное проявление признаков аллелей.
Кодоминирование – независимое проявление аллелей в гетерозиготе.
В некоторых случаях механизм взаимодействия аллелей расшифрован. Лучше всего их взаимоотношения иллюстрируют белки-ферменты. Данные биохимической генетики показывают, что явление доминирования часто связано с активностью определенного фермента.
Менее понятно явление моногенного гетерозиса, когда в гетерозиготе признак проявляется сильнее, чем в гомозиготе по доминантному аллелю. Гетерозис – явление превосходства гибридов над обеими родительскими формами. Феномен гетерозиса интенсивно изучался в связи с его важной ролью для селекции. Это явление весьма сложное и неоднозначное. Неоднозначно понятие "превосходство", так как гетерозис проявляется на репродуктивном, соматическом и адаптивном уровнях. Проявления на разных уровнях могут быть противоположной направленности (например, "улучшение" соматических показателей может сопровождаться "ухудшением" адаптивных). Неоднозначны сами понятия "улучшение", "ухудшение", поскольку они не имеют четких критериев и их применение крайне субъективно. Существует несколько теорий, объясняющих природу гетерозиса.
Феномен моногенного гетерозиса, когда показана зависимость признака от одной аллельной пары, является только одним аспектом теории гетерозиса. По предложению Ф. Добжанского это явление получило название сверхдоминирования. Оно имеет большое значение для эволюционной теории, поскольку демонстрирует преимущество гетерозигот в популяциях. Однако это понятие скорее применимо к адаптивному уровню, поэтому до выяснения генетических механизмов сверхдоминирования рассматривать его как особый вид межаллельных взаимодействий преждевременно.
В живых организмах часто взаимодействуют не только аллели одного гена, но и аллели разных генов, давая самые различные варианты расщепления. Различают три основных типа взаимодействия неаллельных генов.
Комплементарность – взаимодействие разных доминантных аллелей обусловливает появление нового признака.