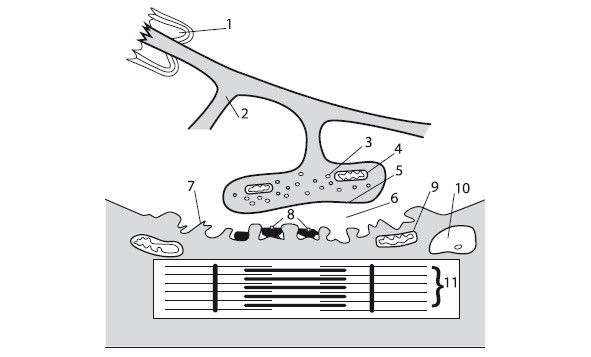
Рис. 1.4. Схема структуры нервно-мышечного синапса 1 – миелиновая оболочка аксона; 2 – концевые веточки аксона; 3 – пузырьки, содержащие ацетилхолин; 4 – митохондрия; 5 – пресинаптическая мембрана, покрывающая концевую веточку аксона в зоне нервно-мышечного синапса; 6 – синаптическая щель; 7 – постсинаптическая мембрана, покрывающая мышечное волокно в зоне нервно-мышечного синапса; 8 – ацетилхолинорецепторы на постсинаптической мембране; 9 – митохондрия мышечного волокна; 10 – ядро мышечной клетки; 11 – миофибрилла
В пресинаптической части нервно-мышечного соединения вырабатывается и собирается в особые пузырьки химический передатчик возбуждения – ацетилхолин (АХ). В механизме передачи возбуждения от нерва на мышечные волокна выделяют три последовательных процесса: электрохимический, включающий деполяризацию концевой веточки аксона и выделение ацетил-холина в синаптическую щель; химический, основу которого составляет диффузия медиатора АХ к постсинаптической мембране и образование на ней его комплекса с холинорецептором; электрический, включающий увеличение ионной проницаемости постсинаптической мембраны, возникновение локального электрического потенциала (потенциала концевой пластинки), развитие потенциала действия мышечного волокна.
Действие выброшенного в синаптическую щель АХ краткосрочно. Временно возникающий на постсинаптической мембране комплекс "АХ-рецептор" после прохождения каждого импульса разрушается ферментом ацетилхолинэстеразой. Запасов АХ в нервном окончании достаточно для проведения примерно 10 000 импульсов. При длительной импульсации мотонейронов, несмотря на постоянный синтез АХ, его содержание в концевых веточках может постепенно уменьшаться. В результате этого возможны нарушения передачи возбуждения в нервно-мышечных синапсах – пресинаптический нервно-мышечный блок. В процессе напряженной и длительной мышечной работы АХ может не успевать разрушаться и накапливается в синаптической щели. Способность постсинаптической мембраны к генерации потенциала концевой пластинки при этом уменьшается, и развивается частичный или полный постсинаптический нервно-мышечный блок. В этом случае сократительные свойства мышечных волокон снижаются.
Передача возбуждения от одного нейрона на другой происходит аналогичным образом через синапс между двумя нейронами. Так же как и в нервно-мышечном синапсе, медиатор поступает в синаптическую щель и активирует постсинаптическую мембрану дендритов или тело другого нейрона.
Сегментарная регуляция мышечной активности. Нейроны спинного мозга имеют довольно сложную организацию. В передних рогах спинного мозга мотонейроны располагаются упорядоченно и сгруппированы в группы (пулы) в соответствии с принадлежностью к иннервации определенных мышц туловища и конечностей. Это позволяет избирательно активировать ту или иную группу мышц. В передних рогах спинного мозга также находятся интернейроны, которые обеспечивают взаимодействие между нейронами, как на уровне одного сегмента, так и межсегментарно. Именно сегментарные связи нейронов являются основой для сегментарных рефлексов – сухожильных. На базе межсегментарных связей реализуются спинальные рефлексы: сгибательный, чесательный, шаговый.
Все нейроны серого вещества спинного мозга располагаются в определенном порядке. Наиболее общепринятым является цитоархитектоническое подразделение серого вещества на пластинки Рекседа, которые нумеруются в дорсовентральном направлении (рис. 1.5).
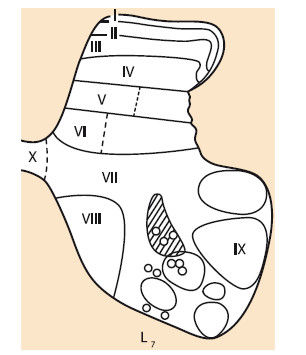
Рис. 1.5. Схема разделения серого вещества спинного мозга 7-го поясничного сегмента кошки и локализация клеток Реншоу и тормозных нейронов системы 1а (В.И. Сафьянц, 1976)
I-Х – пластины серого вещества; кружки – клетки Реншоу; заштрихованная часть – тормозные нейроны системы 1а
Пластины I и III образованы желатинозной субстанцией Роланда. Желатинозный комплекс регулирует синаптическое проведение сигналов от первичных афферентов и супраспинальных структур на нейронные ансамбли спинного мозга. Нейроны желатинозной субстанции активируются высокопороговыми кожными и мышечными афферентами и тормозятся низкопороговыми.
Пластины I–III, IV квалифицируются как первичная сенсорная область серого вещества, в нее проецируется большая часть афферентных волокон от туловища и конечностей.
Интернейроны, составляющие пластины V и VI, связаны с высокопороговыми афферентами сгибательного рефлекса, а также низкопороговыми афферентами мышц туловища и конечностей. Пластины VII и VIII содержат интернейроны с разнообразными афферентными входами. В вентральном отделе VIII пластины располагаются клетки Реншоу.
Интернейроны спинного мозга различают также в зависимости от того, какими надсегментарными структурами и нисходящими путями регулируется их активность. Так, интернейроны, связанные с корково-спинномозговыми (пирамидными) нейронами, располагаются в латеральных отделах V и VI пластин, с красноядерно-спинномозговыми волокнами – в латеральных отделах VI и VII, с преддверно-спинномозговыми и ретикулярно-спинномозговыми – в медиальных отделах VII и VIII пластин. Каждая функциональная группа интернейронов принимает участие в организации и осуществлении определенного вида целенаправленной деятельности спинного мозга, которая модулируется сигналами от соответствующих рецепторных систем или нисходящими командами от различных структур головного мозга. Именно интернейронная система определяет специфику эффекторной деятельности спинного мозга, активируя необходимые для ее выполнения моторные пулы и составляющие их мотонейроны (К.В. Баев, 1984; Ю.П. Герасименко, 2000; Р.М. Городничев и др., 2012).
Параметры эфферентной возбуждающей и тормозящей импульсации, направленной к мотонейронам, определяют частоту разрядов мотонейрона и, следовательно, характеристики активности скелетных мышц, скорость и силу сокращения их мышечных волокон. Например, для поддержания вертикальной позы требуется относительно небольшая интенсивность возбуждающих влияний, которая обеспечивает активацию лишь низкопороговых медленных ДЕ. В этом случае активированные ДЕ поддерживают небольшое по величине, но практически постоянное и без утомления напряжение мышц. Совершенно другая ситуация наблюдается при выполнении максимальных усилий. Возбуждающие эфферентные влияния приближаются к своему максимуму и обеспечивают активацию как низкопороговых, так и высокопороговых мотонейронов. При этом и медленные и быстрые ДЕ развивают максимальные по силе сокращения своих мышечных волокон.
Характеристики импульсной активности мотонейрона определяются алгебраической суммацией всех возбуждающих и тормозных постсинаптических потенциалов, степенью приближения величины мембранного потенциала к критическому уровню деполяризации, необходимому для возникновения потенциала действия данного мотонейрона. Активность мотонейрона проявляется по-разному: в виде одиночного импульса, серии импульсов, нескольких серий, возникающих эпизодически или повторяющихся с определенной периодичностью.
Характер импульсной активности мотонейрона зависит, с одной стороны, от его собственных свойств – возбудимости и лабильности, а с другой – от параметров эфферентного нисходящего воздействия из структур головного мозга и влияния афферентной импульсации от проприорецепторов мышц. Эти факторы приводят к трансформации ритма импульсации мотонейрона, ее модуляции.
Организация мышечной активности сегментарными структурами спинного мозга довольно сложна. Она включает механизмы автоматизированного и коррекционного включения и выключения мотонейронов, обычно на уровне нескольких сегментов. Набор спинальных двигательных автоматизмов относительно невелик. По своей природе они, в основном, являются врожденными. В процессе онтогенеза интернейроны, ответственные за реализацию спинальных автоматизмов, попадают под все более эффективный регулирующий контроль надсегментарных нисходящих влияний. Постепенно спинномозговые рефлекторные автоматизмы редуцируются и уступают место более высокоорганизованным двигательным реакциям, регуляция которых осуществляется над-сегментарными структурами. В настоящее время различают следующие рефлекторные автоматизмы, обеспечивающиеся целиком нейрональными структурами спинного мозга.
Рефлекс на растяжение проявляется в рефлекторном сокращении мышцы в ответ на ее растягивание. Рецепторным аппаратом этого рефлекса является совокупность мышечных веретен. Реализуется рефлекс через моносинаптическую рефлекторную дугу и сопровождается реципрокным торможением мышцы-антагониста за счет активации коллатералями афферентных волокон группы тормозящих интернейронов. Рефлекс на растяжение имеет быстрый (фазический) компонент и более слабый и длительный тонический. Фазический компонент обусловлен активацией афферентами на крупных а-мотонейронов, а тонический – активацией вторичными нервными окончаниями (II) малых а-мотонейронов. В основе всех сухожильных рефлексов лежит рефлекс на растяжение. Обычные сухожильные рефлексы являются фазическими. Мышечный тонус, довольно часто определяемый в практике спортивных исследований, по своей природе является отражением тонического рефлекса на растяжение.