Если каждый последующий нервный импульс приходит к мышечному волокну в фазу его расслабления, то происходит суммирование (суперпозиция) механических ответов. При непрерывной импульсации мотонейрона иннервируемая им группа мышечных волокон будет все время развивать силу, но величина этой силы будет колебаться, так как после каждого очередного импульса мышечные волокна успевают частично расслабиться (рис. 2.4). Этот режим сокращения и называется зубчатым тетанусом. Он характерен, например, для работы мышц конечностей при поддержании вертикальной позы.
Если частота импульсации мотонейронов столь высока, что выходящий из саркоплазматического ретикулума кальций не успевает вновь вернуться за слишком короткие промежутки между смежными нервными импульсами, мышечные волокна будут находиться в состоянии максимального напряжения. В этих условиях каждый очередной нервный импульс приходит к мышечному волокну в фазу подъема напряжения. При этом будет поддерживаться постоянное, максимальное для мышечных волокон усилие (рис. 2.4). Такой режим сокращения называется гладким тетанусом. ДЕ работают в режиме гладкого тетанического сокращения при развитии произвольных максимальных усилий.
Сравнение одиночного мышечного сокращения с тетаническим показывает, что развиваемое при гладком тетанусе напряжение в 2–4 раза больше, чем при одиночном сокращении. Напряжение при тетаническом сокращении по сравнению с одиночным возрастает в связи с тем, что последовательно расположенные упругие элементы мышцы (сухожилия мышцы, участки саркомеров, места перехода миофибрилл в соединительную ткань) успевают растянуться под влиянием развивающегося напряжения до максимума. Этого не происходит при кратковременном одиночном сокращении. Кроме того, при тетаническом сокращении создаются возможности для прикрепления большего числа поперечных мостиков. Это также приводит к повышению развиваемого усилия. Гладкий тетанус для быстрых и медленных ДЕ достигается при разной частоте импульсации. Это связано с различной длительностью их одиночного сокращения. Так, например, гладкое тетаническое сокращение у медленных ДЕ камбаловидной мышцы достигается при частоте 30 имп/с, а у быстрых ДЕ глазодвигательных мышц – при 350 имп/с.
В режиме тетанического сокращения мышца способна работать лишь непродолжительное время. Причиной этого является недостаточное расслабление мышцы и неполное восстановление ее энергетического потенциала, израсходованного в процессе развития напряжения мышечных волокон. Наиболее утомителен режим гладкого тетануса. Сократительные свойства мышцы во многом зависят от предыстории ее активности. Так, например, отдельный стимул сразу после расслабления мышцы из состояния тетануса вызывает гораздо более сильное одиночное сокращение, чем в обычных условиях. Увеличение силы или амплитуды одиночного ответа мышцы под влиянием ее предварительной активности называется потенциацией. Степень потенциации у разных мышц неодинакова. Причины потенциации известны пока плохо. По-видимому, предшествующая активность вызывает изменения в системе выброса ионов кальция в тропонин-тропомиозиновом регуляторном комплексе и в самих сократительных белках – актине и миозине. Наличие феномена потенциации наряду с другими факторами определяют высокую вариативность сократительных свойств мышц человека.
Зависимость между исходной длиной и усилием, развиваемым скелетной мышцей. Кроме последовательно расположенных упругих компонентов в мышце также имеются и параллельные упругие компоненты – соединительно-тканные образования, составляющие оболочку мышечных волокон и их пучков. Последовательные и упругие компоненты во многом определяют механические свойства скелетных мышц.
Покоящаяся мышца обладает упругостью. Это проявляется в развитии напряжения мышцы при ее растягивании. Чем больше растягивается мышца, тем больше в известных пределах развиваемое ею напряжение. При отсутствии какой-либо внешней нагрузки мышца принимает равновесную длину. В таком положении ее упругое напряжение равно нулю. В живом организме человека длина мышцы всегда превышает равновесную и, следовательно, даже расслабленные мышцы имеют некоторое напряжение.
Сила, развиваемая мышцей при произвольном максимальном сокращении или при искусственном электрическом раздражении, зависит от исходной длины мышцы (рис. 2.5).
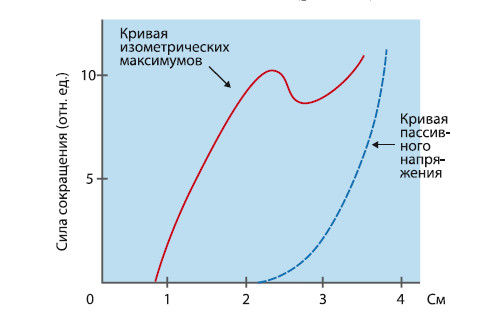
Рис. 2.5. Зависимость напряжения мышцы от ее исходной длины
Прирост силы во время сокращения суммируется с пассивным напряжением упругих компонентов мышцы. Напряжение сокращающейся мышцы будет максимальным, когда ее длина составляет около 120 % равновесной.
Длину мышцы, при которой развиваемое ею напряжение максимально, называют длиной покоя. При более значительном предварительном растягивании напряжение при сокращении снижается. Так, при длине, составляющей 180 % равновесной, оно достигает около 20 % от максимального. Напряжение понижается и в том случае, если ее длина меньше равновесной. В этих условиях возникающее при сокращении напряжение будет тем меньше, чем при меньшей исходной длине сокращается мышца. В обычных условиях длина покоя у многих скелетных мышц человека достигается при крайней или близкой к ней степени растяжения. Следовательно, у большинства мышц напряжение будет максимальным, когда они максимально растянуты, а костные рычаги, к которым прикреплена данная мышца, находятся на предельно возможном расстоянии друг от друга.
Характер зависимости величины напряжения, развиваемого мышцей, от ее исходной длины определяется условиями взаимодействия сократительных миозиновых и актиновых нитей. Как указывалось ранее, развиваемое мышцей напряжение зависит от числа активных поперечных мостиков, осуществляющих втягивание актиновых нитей в промежутки между миозиновыми миофиламентами. Оптимальные условия для активации наибольшего числа поперечных мостиков создаются при длине покоя. При меньшей длине мышцы число активных поперечных мостиков уменьшается. К тому же укорочение мышцы приводит к некоторому нарушению проведения волны возбуждения по Т-системе и процесса освобождения ионов кальция из саркоплазматического ретикулума. В результате этих причин величина развиваемого напряжения снижается. Если мышца растянута больше по сравнению с длиной покоя, то напряжение также уменьшится, потому что актиновые нити вытягиваются из миозиновых. Так, при длине мышцы, равной 145 % ее длины покоя, развивается напряжение примерно в 50 % от максимума, поскольку зона перекрытия актиновых и миозиновых нитей составляет лишь половину возможной, и только половина поперечных мостиков может прикрепляться к актиновым нитям. При крайней степени растяжения мышцы сверх длины покоя напряжение не развивается, потому что актиновые и миозиновые нити не перекрываются, т. е. условия для активации поперечных мостиков вовсе отсутствуют.
Зависимость между силой и скоростью сокращения мышцы.
Кривая зависимости между силой и скоростью концентрического сокращения мышцы представлена на рис. 2.6. Как видно на графике, чем больше скорость сокращения, тем меньше развиваемая мышцей сила. Такая зависимость наблюдается как в опытах на изолированной мышце, так и при выполнении движений в естественных условиях. Скорость концентрического сокращения зависит от величины внешней нагрузки: величина укорочения в единицу времени тем больше, чем меньше нагрузка.
Максимальная скорость наблюдается у свободной от нагрузки мышцы. Если нагрузка равняется величине изометрической силы, мышца вообще не укорачивается. В этом случае она развивает напряжение, соответствующее ее максимальному изометрическому напряжению. Поэтому максимальная мышечная сила развивается при медленных движениях, например, когда поднимается тяжелый вес.
Характер зависимости между силой и скоростью мышечного сокращения определяется особенностями взаимодействия миозиновых и актиновых нитей при быстрых и медленных движениях.
В процессе быстрого укорочения мышцы актиновые и миозиновые нити взаимодействуют очень непродолжительное время. Поэтому активируется меньшее число поперечных мостиков, и мышца развивает меньшее напряжение, чем при медленном укорочении. С какой скоростью ни укорачивалась бы мышца при концентрическом сокращении, развиваемое в этом случае напряжение не достигает величины, регистрируемой при изометрическом сокращении.
При эксцентрическом сокращении (удлинении сокращающейся мышцы) развивается большее напряжение, чем в условиях изометрии. С увеличением скорости удлинения мышцы величина максимума напряжения возрастает.